04/01/2024
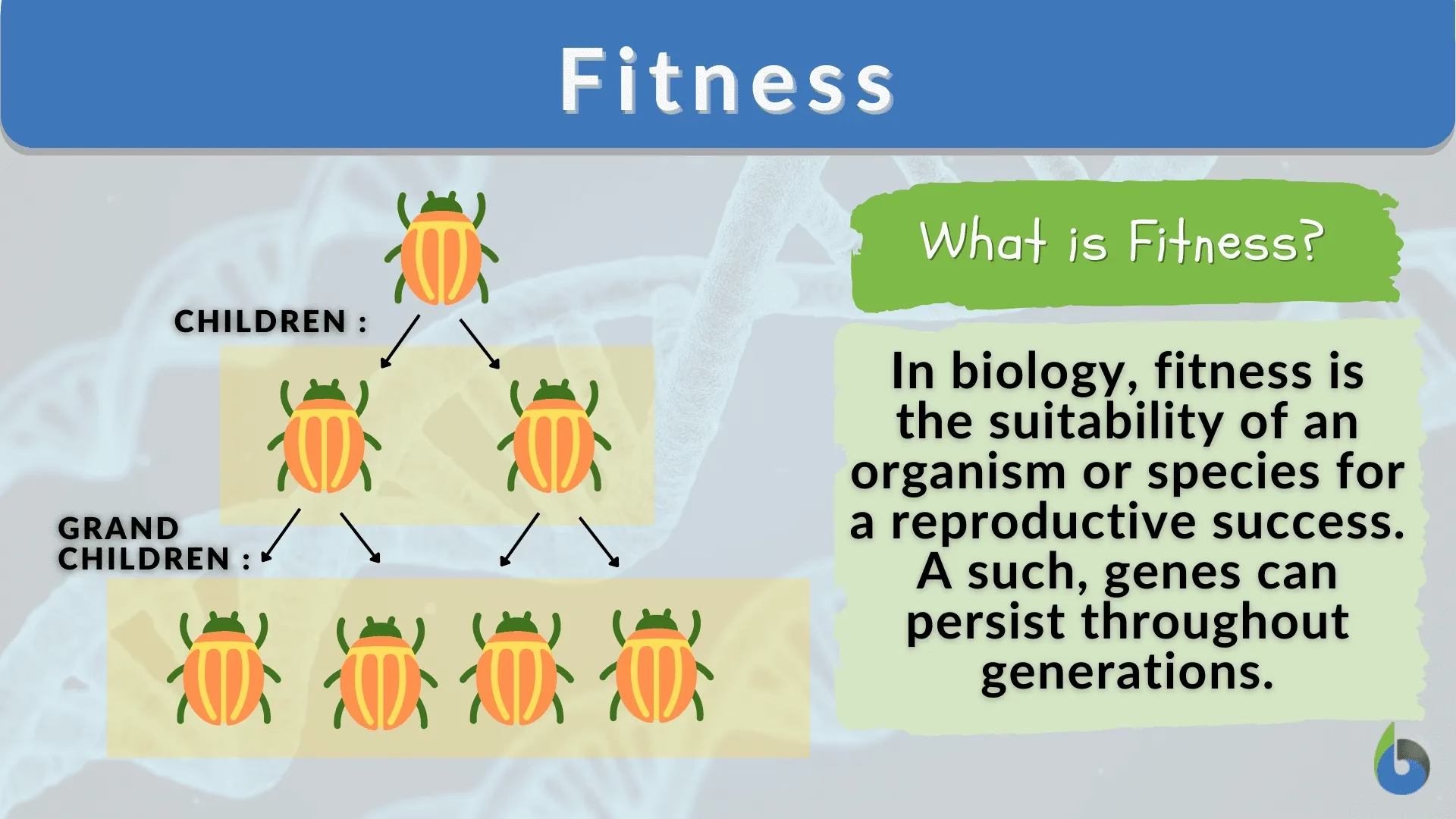
În lumea fitness-ului, adesea ne gândim la mușchi sculptați, anduranță și sănătate vibrantă. Dar în domeniul vast și complex al biologiei evoluționiste, termenul „fitness” capătă o semnificație cu totul diferită, una fundamentală pentru înțelegerea modului în care viața evoluează. Nu este vorba despre cât de repede alergi sau cât de mult ridici greutăți, ci despre capacitatea supremă a unui organism de a se reproduce și de a-și transmite genele generațiilor următoare. Această noțiune, adesea contraintuitivă pentru publicul larg, este piatra de temelie a selecției naturale și a proceselor care modelează diversitatea uimitoare a speciilor de pe Pământ. Este esențial să înțelegem că, în acest context științific, „fitness” nu se referă la condiția fizică individuală, ci la succesul reproductiv al unui genotip într-un anumit mediu. Un individ care trăiește mult, dar nu are urmași, are un fitness evoluționist zero, indiferent de forța sau sănătatea sa.
Conceptul de fitness, deși nu a fost „inventat” de o singură persoană în sensul modern al geneticii, a evoluat odată cu teoria evoluției. Charles Darwin a recunoscut importanța variabilității ereditare și a „supraviețuirii celui mai apt” (termen popularizat de Herbert Spencer), dar mecanismul precis al moștenirii i-a scăpat. În cele ce urmează, vom explora în detaliu ce înseamnă fitness-ul în genetica modernă, cum este măsurat și cum a fost înțeleasă această idee centrală de-a lungul istoriei științei.
Ce Înseamnă „Fitness” în Genetică?
În genetică, „fitness-ul” nu este pur și simplu numărul real de urmași produși de un individ, ci mai degrabă o propensitate sau o probabilitate. Această distincție este crucială. De exemplu, așa cum a subliniat Maynard Smith, „fitness-ul este o proprietate, nu a unui individ, ci a unei clase de indivizi – de exemplu, homozigoți pentru alela A la un anumit locus. Astfel, expresia „numărul așteptat de urmași” înseamnă numărul mediu, nu numărul produs de un singur individ. Dacă primul sugar uman cu o genă pentru levitație ar fi fost lovit de un fulger în cărucior, acest lucru nu ar dovedi că noul genotip are un fitness scăzut, ci doar că acel copil a fost ghinionist.” Această perspectivă subliniază că fitness-ul este o măsură probabilistică a succesului reproductiv pe termen lung al unui anumit genotip, în contextul condițiilor de mediu și al interacțiunilor cu alți indivizi.
Practic, fitness-ul unui individ – având un anumit set de fenotipuri – este probabilitatea ca acel individ să fie inclus în grupul selectat ca părinți ai generației următoare. Pentru a înțelege mai bine acest concept, biologii evoluționiști utilizează două operaționalizări comune ale fitness-ului, în special în contextul populațiilor asexuate, fără recombinare genetică, unde fitness-urile pot fi atribuite direct genotipurilor.
Modele de Fitness: Absolut vs. Relativ
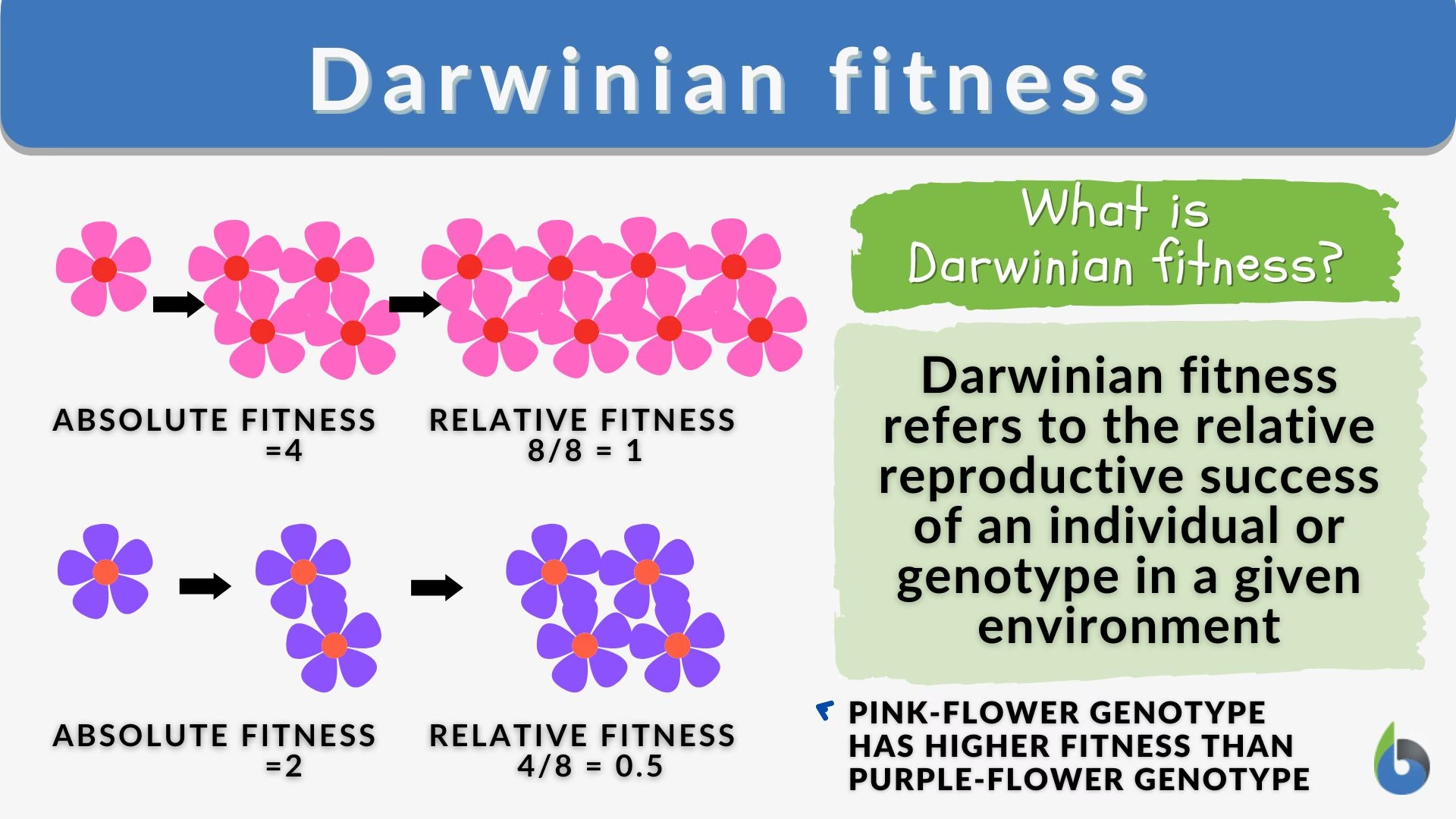
Fitnessul Absolut (W)
Fitnessul absolut (W) al unui genotip este definit ca modificarea proporțională a abundenței acelui genotip de-a lungul unei generații, atribuită selecției. Într-o populație infinit de mare, unde nu există deriva genetică și neglijând schimbările de abundență datorate mutațiilor, relația este simplă: n(t+1) = W * n(t), unde n(t) este abundența genotipului la generația t. Un fitness absolut mai mare de 1 indică o creștere a abundenței genotipului, în timp ce un fitness absolut mai mic de 1 indică un declin. De exemplu, dacă o colonie de bacterii cu un anumit genotip are un W = 2, înseamnă că populația de bacterii cu acel genotip se va dubla la fiecare generație. Dacă W = 0.5, populația se va înjumătăți. Acest model este util pentru a înțelege dinamica populațiilor în termeni absoluți, oferind o imagine clară a creșterii sau descreșterii numerice a unui anumit genotip.
Fitnessul Relativ (w)
În timp ce fitness-ul absolut determină modificările în abundența genotipului, fitness-ul relativ (w) determină modificările în frecvența genotipului. Dacă N(t) este dimensiunea totală a populației la generația t, iar frecvența genotipului relevant este p(t) = n(t) / N(t), atunci p(t+1) = (w / w_bar) * p(t), unde w_bar este fitness-ul relativ mediu în populație. Fitness-urile relative indică doar schimbarea prevalenței diferitelor genotipuri unele față de altele, deci valorile lor relative sunt cele importante. Ele pot fi orice număr non-negativ, inclusiv 0. Este adesea convenabil să alegem un genotip ca referință și să îi setăm fitness-ul relativ la 1. Fitness-ul relativ este utilizat în modelele standard Wright-Fisher și Moran de genetică a populațiilor, care se concentrează pe evoluția frecvențelor alelice și genotipice.
Este important de reținut că fitness-urile absolute pot fi folosite pentru a calcula fitness-ul relativ, deoarece p(t+1) = n(t+1) / N(t+1) = (W / W_bar) * p(t), unde W_bar este fitness-ul absolut mediu în populație. Aceasta implică faptul că w / w_bar = W / W_bar, sau, cu alte cuvinte, fitness-ul relativ este proporțional cu W / W_bar. Cu toate acestea, nu este posibil să se calculeze fitness-urile absolute doar din fitness-urile relative, deoarece fitness-urile relative nu conțin informații despre modificările abundenței totale a populației N(t). Atribuirea valorilor de fitness relativ genotipurilor este adecvată matematic atunci când sunt îndeplinite două condiții: în primul rând, populația se află în echilibru demografic și, în al doilea rând, indivizii variază în rata lor de naștere, capacitatea de competiție sau rata de deces, dar nu o combinație a acestor trăsături. Alegerea modelului de fitness depinde de întrebarea specifică la care se încearcă să se răspundă și de datele disponibile.
Tabel Comparativ: Fitness Absolut vs. Relativ
| Caracteristică | Fitness Absolut (W) | Fitness Relativ (w) |
|---|---|---|
| Definește | Modificarea proporțională a abundenței genotipului în timp | Modificarea frecvenței genotipului în raport cu alte genotipuri |
| Valoare | Poate fi orice număr pozitiv. W > 1 (creștere), W < 1 (scădere), W = 1 (stabilitate) | Poate fi orice număr non-negativ. Valorile sunt importante doar relativ una la alta (adesea un genotip de referință este setat la 1) |
| Informații despre populație | Reflectă creșterea/scăderea numerică a populației totale | Nu oferă informații despre dimensiunea totală a populației, ci doar despre compoziția sa relativă |
| Utilizare tipică | Analiza creșterii sau declinului populaționale, demografie | Modele de genetică a populațiilor (Wright-Fisher, Moran) pentru a urmări schimbările de frecvență ale alelelor |
| Calcul | Necesită numărul de indivizi la generații succesive | Calculat din fitness-ul absolut prin normalizare față de fitness-ul mediu al populației |
Schimbarea Frecvențelor Genotipului Datorită Selecției
Modificarea frecvențelor genotipului datorită selecției decurge direct din definiția fitness-ului relativ: Δp = p(t+1) - p(t) = ((w - w_bar) / w_bar) * p(t). Astfel, frecvența unui genotip va scădea sau va crește în funcție de faptul dacă fitness-ul său este mai mic sau mai mare decât fitness-ul mediu al populației, respectiv. Într-o populație, genotipurile cu fitness mai mare vor deveni mai comune în timp, în timp ce cele cu fitness mai mic vor scădea în frecvență, putând chiar dispărea.
În cazul particular în care există doar două genotipuri de interes (de exemplu, reprezentând invazia unei noi alele mutante), modificarea frecvențelor genotipului este adesea scrisă într-o formă diferită. Să presupunem că două genotipuri, A și B, au fitness-uri wA și wB, și frecvențe p și 1 - p, respectiv. Atunci fitness-ul mediu este w_bar = wA*p + wB*(1-p). Modificarea frecvenței genotipului A este Δp = ((wA - wB) / w_bar) * p(1-p). Astfel, schimbarea frecvenței genotipului A depinde crucial de diferența dintre fitness-ul său și fitness-ul genotipului B.

Dacă A este mai „fit” decât B, putem defini coeficientul de selecție s prin wA = (1 + s)wB. Coeficientul s măsoară avantajul selectiv al genotipului A. Atunci, Δp = (s / (1 + sp)) * p(1-p), care, pentru valori mici ale lui s (s << 1), poate fi aproximată la Δp ≈ s * p(1-p). Aceasta înseamnă că frecvența genotipului mai „fit” crește aproximativ logistic. Chiar și un coeficient de selecție mic poate duce la schimbări semnificative în compoziția genetică a unei populații pe parcursul multor generații, demonstrând puterea constantă a selecției naturale.
Încărcătura Genetică: Costul Adaptării
Conceptul de încărcătură genetică (Genetic load) măsoară fitness-ul mediu al unei populații de indivizi, relativ fie la un genotip teoretic de fitness optim, fie la cel mai „fit” genotip prezent efectiv în populație. Dacă avem n genotipuri A1...An, cu fitness-uri w1...wn și frecvențe genotipice p1...pn, încărcătura genetică (L) poate fi calculată ca: L = (w_max - w_bar) / w_max, unde w_max este fitness-ul maxim observat sau teoretic, iar w_bar este fitness-ul mediu al populației.
Încărcătura genetică poate crește atunci când mutațiile dăunătoare (deleterii), migrația, consangvinizarea (înrudirea) sau încrucișările între populații diferite scad fitness-ul mediu. De exemplu, consangvinizarea crește probabilitatea ca indivizii să moștenească două copii ale unei alele recesive dăunătoare, scăzând fitness-ul mediu al populației. Încărcătura genetică poate crește și atunci când mutațiile benefice cresc fitness-ul maxim față de care sunt comparate alte mutații; aceasta este cunoscută sub numele de încărcătură de substituție sau costul selecției. Este un concept important care ne ajută să înțelegem presiunile evolutive și compromisurile energetice și genetice pe care le suportă populațiile.
Moștenirea și Darwin: O Piesă Lipsă în Puzzle-ul Evoluției
Teoria selecției naturale a lui Charles Darwin, deși revoluționară, a fost incompletă din punct de vedere logic din cauza lipsei unei explicații adecvate a eredității. Darwin a înțeles profund că evoluția depinde de existența variabilității ereditare în cadrul unei specii pentru a genera diferențele dintre populațiile ancestrale și cele descendente. În „Originea Speciilor”, el a dedicat capitole întregi documentării variabilității și eficacității selecției artificiale. Cu toate acestea, el s-a confruntat cu o dificultate majoră legată de modul în care trăsăturile erau transmise de la o generație la alta.
Darwin a fost influențat de ideea predominantă a „moștenirii prin amestec” (blending inheritance), conform căreia caracteristicile părinților se amestecă în urmași, la fel cum se amestecă culorile. Această idee prezenta o problemă serioasă: sub moștenirea prin amestec, variația genetică într-o populație ar trebui să se diminueze rapid, deoarece fiecare generație ar deveni mai uniformă, iar genotipurile urmașilor ar fi intermediare între cele ale părinților. Acest lucru ar fi subminat capacitatea selecției naturale de a produce schimbări permanente și cumulative într-o populație, deoarece variația, materia primă a selecției, ar fi fost constant epuizată. Prin urmare, Darwin a simțit nevoia unui mecanism care să genereze continuu variabilitate, pentru a compensa această „diluare”.
Modelul „Pangenesis” al lui Darwin
Pentru a depăși această dificultate și pentru a explica sursa abundentă de variabilitate necesară selecției, Darwin a propus propria sa teorie a moștenirii, numită „pangenesis”, în lucrarea sa „Variația animalelor și plantelor sub domesticire” (1868). Conform acestei ipoteze, fiecare parte a corpului unui organism producea mici particule ereditare, numite „gemule”, care erau colectate în celulele sexuale și transmise urmașilor. Această teorie implica moștenirea caracterelor dobândite în timpul vieții unui organism – o idee cunoscută și sub numele de lamarckism, deși Darwin a avut o viziune mai nuanțată decât Jean-Baptiste Lamarck. Prin gemule, variațiile experimentate de individ de-a lungul vieții sale (cum ar fi dezvoltarea mușchilor prin exerciții sau adaptările la un mediu specific) ar fi fost transmise celulelor germinale și, ulterior, urmașilor. Pangenesis oferea un mecanism prin care variația putea fi generată și transmisă, rezolvând aparent problema epuizării variației sub moștenirea prin amestec.
Cu toate acestea, Darwin însuși a recunoscut că pangenesis nu putea explica toate situațiile evoluționiste. Un exemplu notabil, pe care l-a folosit pentru a contrazice ideile lamarckiene, a fost cazul castelor sterile de insecte sociale. Aceste insecte (furnici, albine) prezintă adaptări complexe, dar nu se reproduc. Cum ar fi putut „gemulele” lor acumula adaptări și le transmite mai departe, dacă ele însele nu aveau urmași? „Nicio cantitate de exercițiu, sau obișnuință, sau voință, la membrii absolut sterili ai unei comunități nu ar fi putut afecta structura sau instinctele membrilor fertili, care singuri lasă descendenți,” a scris Darwin. Aceasta a fost o dovadă puternică împotriva moștenirii caracterelor dobândite, arătând că teoria sa avea limitări semnificative.
Mendel la Salvare: Moștenirea Particulată
Tragic, Darwin nu a fost conștient de munca lui Gregor Mendel, publicată cu doi ani înainte de „Variația animalelor și plantelor sub domesticire”. Spre deosebire de Darwin, Mendel a rezolvat problema moștenirii prin utilizarea unei abordări unice: combinarea experimentelor genetice riguroase cu predicții cantitative, probabilistice. Mendel a propus ipoteza moștenirii particulate, potrivit căreia trăsăturile sunt transmise prin unități discrete (gene), care își păstrează integritatea de-a lungul generațiilor, fără a se amesteca. Această descoperire monumentală a eliminat dificultățile puse de moștenirea prin amestec. Variația într-o populație este conservată, nu pierdută, atunci când nu acționează forțe evolutive, un echivalent genetic al legii inerției a lui Galileo. Astfel, nu mai era nevoie de un mecanism constant de generare a variației, deoarece variația existentă era pur și simplu redistribuită, nu diluată.
Deși Darwin a analizat date din propriile sale experimente de încrucișare (de exemplu, cu speciile de Primula) care prezentau, retrospectiv, raporturi mendeliene clare, el nu a reușit să înțeleagă semnificația acestor rezultate, deoarece nu avea un model de moștenire particulară aplicabil datelor genetice. Abordarea sa, deși adesea cantitativă, nu a atins nivelul de rigoare teoretică a lui Mendel. Redescoperirea lucrării lui Mendel la începutul secolului XX a revoluționat înțelegerea eredității și, în cele din urmă, a permis o sinteză între genetică și evoluție.
Scepticismul față de selecția naturală a persistat mult timp, până în anii 1920, parțial din cauza lipsei de dovezi directe și a neînțelegerii mecanismului eredității. Proeminenți geneticieni timpurii, precum William Bateson și Hugo de Vries, erau sceptici față de rolul selecției naturale, punând accentul pe mutații cu efecte mari. Odată ce baza particulară a geneticii a fost înțeleasă, s-a văzut că aceasta permitea variației să fie transmisă intactă noilor generații, iar evoluția putea fi apoi înțeleasă ca un proces de schimbare a frecvențelor variantelor stabile. Astfel, genetica evoluționistă s-a dezvoltat ca o parte centrală a biologiei, confirmând și extinzând principiile darwiniene, oferind un cadru solid pentru înțelegerea mecanismelor profunde ale vieții.
Întrebări Frecvente (FAQ)
1. Este „fitness” evoluționist același lucru cu puterea fizică sau sănătatea?
Categoric nu. În biologia evoluționistă, „fitness-ul” este definit strict prin succesul reproductiv. Un individ poate fi extrem de puternic și sănătos, dar dacă nu se reproduce sau produce urmași care nu supraviețuiesc până la vârsta reproductivă, fitness-ul său este zero. În contrast, un organism fragil, dar care produce un număr mare de urmași viabili, care la rândul lor se reproduc, are un fitness evoluționist ridicat. Este o măsură a contribuției genetice la generația următoare, nu a vigorii individuale.
2. Cum se măsoară „fitness-ul” în practică?
Măsurarea directă a fitness-ului poate fi dificilă în natură, deoarece necesită urmărirea succesului reproductiv al indivizilor de-a lungul întregii lor vieți și pe parcursul mai multor generații. Adesea, este estimată prin componentele sale, cum ar fi rata de supraviețuire (viabilitatea), rata de fertilitate (fecunditatea) și succesul în împerechere. Biologii observă populațiile de-a lungul mai multor generații, urmăresc numărul de urmași viabili produși de diferite genotipuri sau fenotipuri și calculează ratele de creștere sau de declin relativ. Studiile pe termen lung și experimentele de selecție controlată în laborator oferă cele mai precise estimări ale fitness-ului.
3. De ce este important „fitness-ul” în evoluție?
Fitness-ul este motorul principal al selecției naturale. Diferențele de fitness între indivizi sau genotipuri duc la schimbări în frecvențele alelelor și genotipurilor în populație de-a lungul timpului. Organismele cu un fitness mai mare transmit mai multe copii ale genelor lor, ceea ce duce la o adaptare progresivă a populațiilor la mediul lor. Fără variații în fitness, nu ar exista selecție naturală și, prin urmare, nici adaptare evolutivă. Este mecanismul fundamental prin care speciile se modifică și se diversifică.
4. Ce este coeficientul de selecție (s)?
Coeficientul de selecție (s) este o măsură a dezavantajului sau avantajului relativ al unui genotip față de un altul. De exemplu, dacă un genotip are fitness-ul wA și un genotip de referință are fitness-ul wB, iar wA = (1 + s)wB, atunci s reprezintă măsura în care genotipul A este mai „fit” decât B. Un s pozitiv înseamnă un avantaj selectiv, un s negativ un dezavantaj. Chiar și valori mici ale lui s (de exemplu, 0.01) pot avea un impact enorm asupra frecvențelor genelor pe parcursul a sute sau mii de generații, demonstrând cum mici diferențe de fitness pot modela profund evoluția și pot duce la adaptări semnificative.
5. Ce este încărcătura genetică?
Încărcătura genetică se referă la reducerea fitness-ului mediu al unei populații din cauza prezenței alelelor dăunătoare sau a altor factori genetici care împiedică atingerea fitness-ului maxim posibil. Este un concept care reflectă „costul” menținerii variației genetice sau al adaptării la noi condiții. Mutațiile dăunătoare (care reduc funcționalitatea), consangvinizarea (care crește homozigozitatea pentru alele recesive dăunătoare) și anumite tipuri de selecție (cum ar fi selecția de substituție, unde o alelă nouă, benefică, înlocuiește o alelă veche) pot crește încărcătura genetică, indicând o sănătate genetică mai slabă sau un „efort” evolutiv mai mare al populației în ansamblu.
Concluzie
În concluzie, deși termenul „fitness” în limbajul cotidian evocă imagini de forță fizică și sănătate, în biologia evoluționistă, el are o semnificație mult mai profundă și fundamentală: capacitatea unui organism de a se reproduce și de a transmite cu succes materialul său genetic. De la conceptele timpurii ale lui Darwin, care, deși geniale, erau limitate de lipsa unei înțelegeri clare a eredității, până la descoperirile revoluționare ale lui Mendel, care au dezvăluit natura particulate a moștenirii, înțelegerea fitness-ului a evoluat semnificativ. Această interacțiune continuă dintre genetică și studiul evoluției a pavat drumul pentru modele predictive precise și a transformat biologia într-o disciplină mai robustă și mai aplicată.
Principiile darwiniene, îmbogățite de cunoștințele genetice, joacă astăzi un rol mai mare ca niciodată în cercetarea biologică modernă, oferind instrumente valoroase pentru a decodifica complexitatea vieții și a răspunde la întrebări fundamentale despre originea și diversitatea speciilor. Studiile ecologice, genomica populațiilor și cartografierea genelor au demonstrat în mod repetat operația selecției naturale în sălbăticie, confirmând viziunea lui Darwin, dar cu o precizie și o înțelegere moleculară pe care el nu și le-ar fi putut imagina. Așadar, data viitoare când veți auzi cuvântul „fitness”, amintiți-vă că, pe lângă exercițiile fizice, există o dimensiune mult mai vastă, invizibilă, care guvernează viața pe Pământ: succesul implacabil al transmiterii genetice, motorul suprem al evoluției.
Dacă vrei să descoperi și alte articole similare cu Fitnessul în Biologia Evoluționistă: O Cheie a Vieții, poți vizita categoria Fitness.